Replikasi
merupakan proses pengkopian atau duplikasi DNA. Mesin yang mengkopi DNA
memiliki sejumlah protein dan mengikuti aturan-aturan yang ditetapkan untuk
setiap organisme. Replikasi DNA dimulai pada titik khusus yang
disebut Origins of Replication dan berjalan secara dua arah dan
serempak. Proses replikasi DNA secara semikonservatif karena setiap hasil copi
DNA ganda yang baru berisi satu untai (template) yang lama (induk) dan satu
untai komplementer yang baru. DNA Polimerase merupakan enzim yang mengkatalisis
ikatan kovalen antara fosfat dari satu nukleotida dan deoksiribosa (gula) dari
nukleotida berikutnya, DNA polimerase hanya bisa mensintesis untai baru dari arah 5 'ke 3' sehingga arah bacanya dari arah
3' ke 5'.
Terdapat tiga
tahap dalam replikasi DNA yaitu: inisiasi, elongasi dan terminasi.
1. Inisiasi
Terdapat
Primase yang menggabungkan nukleotida RNA menjadi primer. Sebab DNA polimerase
tidak dapat memulai sintesis untai baru,
jadi harus membentuk primer. Kemudian DNA polimerase menambahkan nukleotida DNA
pada primer , DNA polimerase menempel ke ujung 3 'pada RNA primer, DNA lain
mengganti RNA dengan DNA sehingga untai anak lengkap.

2. Elongasi
Pemanjangan DNA
baru dikatakan dengan replikasi garpu. Menggunakan enzim yaitu DNA polimerase.
DNA polimerase menggunakan setiap helai sebagai template pada arah 3' ke 5'.
Jadi DNA baru dapat memanjang hanya pada arah 5' ke 3'. DNA polimerase menambahkan
nukleotida hanya pada ujung untai tumbuh 3'. DNA polymerase III dapat
mensintesis untai komplementer terus menerus dengan memanjangkan DNA yang baru
dengan arah 5' ke 3' dan bergerak menuju
garpu replikasi kemudian menghasilkan leading strand dan lagging strand atau
dapat dikatakan semi-discontinous yaitu leading strands yang bersambung terus-menerus
rantainya dan lagging strands terputus-putus.
Lagging strand
disintesis sebagai rangkaian segmen yang terputus-putus yang disebut fragmen
Okazaki, fragmen Okazaki kemudian akan bergabung dengan DNA ligase membentuk
satu untai DNA tunggal.
Sintesis leading
strands dan lagging strands selama proses replikasi DNA.
1. Pemanjangan
yang dilakukan DNA polimerase III hanya pada arah 5' ke 3'.
2. Salah
satu untai baru, lagging strands, dapat memanjang terus 5' ke 3' ketika
replikasi garpu berlangsung.
3. Untai
baru lainnya, lagging strands harus tumbuh dalam keseluruhan dari arah 3' ke 5'
dengan penambahan
segmen pendek, fragmen Okazaki, yang tumbuh dari atah 5' ke 3'.
4. DNA
ligase bergabung dengan fragmen Okazaki membentuk ikatan bebas antara ujung
mereka menghasilkan untai yang bersambung atau continous.

Pada
leading strands hanya satu primer yang diperlukan untuk sintesis sedangkan
sintesis pada lagging strands , setiap
fragmen Okazaki primer harus secara terpisah.
Enzim yang berperan dalam proses Replikasi DNA
yaitu:
1. Helikase, membuka pilinan DNA heliks ganda,
dimulai dari “origin of
replication”.
2. SSBP, menstabilkan rantai-rantai yang
telah dibuka.
3. Topoisomerases mengurangi puntiran rantai.
4. Primase, membuat RNA primer
yang akan melekat pada rantai template (rantai induk).
5. DNA polymerase memperpanjang rantai
baru dengan menambahkan nukleotida hanya dari ujung 3'.
6.
DNA polimerase I, membuang nukleotida pada RNA primer
dan menggantinya dengan nukleotida DNA.
7.
DNA ligase, menggabungkan ujung 3′ dari DNA yang
menggantikan primer ke bagian lain dari leading strand dan menggabungkan
fragmen-fragmen Okazaki pada laging strand.
A. Proses
replikasi DNA secara keseluruhan
1. DNA Helicase membuka dan memisahkan kedua rantai DNA, dimulai dari origin of replication.

2. Single Stranded Binding Proteins (SSBP) menstabilkan rantai-rantai tunggal dari hasil pemisahan pilinan induk.
1.

3. Topoisomerase mengurangi puntiran
rantai dengan mengatur tingkat pilinan yang disebabkan oleh pemisahan rantai.
4. DNA polymerase hanya dapat menambahkan
nukleotida di ujung 3’ rantai yang ada sebelumnya.
5. DNA Primase membuat RNA primer yang
sesuai dengan rantai template yang akan melekat di rantai template yang akan
menghadirkan rantai dengan ujung bebas 3’ agar DNA polymerase berfungsi.
6. RNA primer dibuat dengan cara
melekatkan ribonucleotida
ke ujung bebas 5’
ribonucleotida sebelumnya.
7. DNA polymerase mensintesis rantai
baru dengan menambah
deoxyribonucleotida ke ujung 3’OH deoxyribonucleotida sebelumnya.
8. Penambahan
deoxyribonucleotida
baru menyebabkan pelepasan molekul
sebuah PPi yang
kemudian dihidrolisis menjadi dua molekul Pi.
9. Rantai
baru selalu disintesis dengan arah orientasi 5’ ke 3’ dan rantai lama dikopi dengan arah 3’ ke 5’.
10. Leading strand direplikasi searah
dengan garpu replikasi dan berlangsung terus menerus.
11. Okazaki fragments merupakan rantai yang
terputus-putus dan mengandung RNA
12. Untuk
menghasilkan rantai DNA yang utuh maka RNA dilepaskan (RNAse H) dan diganti dengan DNA (DNA polymerase) dan
rantai-rantai terputus itu disambung dengan DNA Ligase.

Proses replikasi
prokariot dan eukariot serupa. Perbedaan proses replikasi pada prokariot dan
eukariot yang paling utama adalah berkaitan dengan jumlah DNA. Eukariot
memiliki jumlah DNA jauh lebih banyak dari prokariot (lebih 1000 kali dari
jumlah E. Coli) dan memiliki protein histon dalam nukleosom. Replikasi pada
prokariot terdapat satu titik mulai (origin of replication) sedangkan replikasi
pada eukariot mempunyai banyak titik (origin of replication) tempat awal
replikasi dimulai. DNA polimerase pada eukariot yaitu a,
b, d, g dan e. Sedangkan DNA
polimerase pada prokariot yaitu polimerase I, II, dan III. DNA polimerase pada
prokariotik dan eukariot memiliki fungsi yang berbeda. Pada eukariot fragmen
Okazaki lebih pendek daripada prokariot.

2. PROSES TRANSKRIPSI (PROKARIOT DAN EUKARIOT)
Transkripsi
merupakan proses mensintesis protein gen (serangkaian DNA) dikopi menjadi RNA. Enzim yang digunakan adalah RNA
polimerase yang membuka pilinan kedua untai DNA sehingga terpisah. RNA dapat
menambhakan nukleotida hanya pada ujung 3' dari polimer yang sedang
tumbuh. Molekul RNA memanjang dari arah 5' ke 3'.
Ada
tiga tahapan dalam transkripsi:
1.
Inisiasi – pada tahap ini transcription machinery melekat pada DNA
yang akan dikopi menjadi RNA.
2.
Elongasi – setelah transcription machinery melekat dengan
baik, maka proses transkripsi dimulai dan sebuah rantai RNA baru disintesis.
3.
Terminasi – setelah gen selesai dikopi,
transkripsi dihentikan dan transcription
machinery dan rantai baru RNA melepaskan diri DNA.
A. Proses Transkripsi Eukariot
1. Inisiasi
-
Pada tahap ini, RNA polymerase melekat erat ke DNA
pada poin/urutan basa yang sangat spesifik yang terletak pada at very specific points, Urutan basa
ini disebut promotor.
-
Pada eukariotik, RNA polymerase membutuhkan protein
lain yang disebut transcription factors (TF) untuk memulai transkripsi. Hal ini disebabkan
inisiasi transkripsi pada eukaryotik harus menghadapi masalah pemaketan DNA
yang rumit.
-
Organisme Eukariotik membutuhkan beberapa protein
yang membantu proses inisiasi:
a. Activators – Protein yang melekat ke
tempat khusus DNA untuk menarik RNA
Polymerase. Hal ini penting bagi RNA polymerase untuk mengatasi
masalah pemaketan DNA.
b. Mediators – Protein yang mengizinkan
komunikasi antara activators dan
RNA polymerase dan antara
protein lainnya (other general
transcription factors)
c. Chromatin modifying proteins –
Protein yang memodifikasi paket DNA agar RNA polymerase melekat pada DNA.
-
Begitu polimerase terikat kuat dengan DNA
promotor, kedua untai DNA mengulur dan enzim mulai mentranskripsi untai
cetakannya.
2. Tahap Elongasi
-
Setelah tahap inisiasi dilakukan, RNA polymerase akan bergerak di
sepanjang rantai DNA dan membuka pilinan heliks ganda untuk mengkopinya menjadi
RNA.
-
Polimerase menambahkan nukleotida ke
ujung 3’ dari molekul RNA yang sedang
tumbuh di sepanjang heliks ganda.
-
Pergerakan RNA polymerase di sepanjang rantai
DNA dapat menyebabkan supercoiling.
Hal ini akan diringankan oleh DNA
Topoisomerases
-
Pada eukaryotes, transcription factors
ditukar dengan elongation factors
-
mRNA pada organisme eukaryotic dimodifikasi saat tahap
elongation berlangsung yaitu dengan menambahkan molekul 7-methylguanosine (tutup 5’ atau 5’ cap) ke ujung 5’, untuk menandai mereka sebagai mRNA.
-
Ujung itu ditambah segera setelah proses
DNA dikopi pada 25 Nukleotida pertama. Ujung itu juga memfasilitasi proses
lebih lanjut, export dan translasi mRNA.
-
mRNA
pada eukaryotik mengandung bagian intron dan ekson, bagian intron harus dibuang
karena tidak ikut menentukan urutan asam amino pada protein.
-
Saat RNA polymerase membuat mRNA, intron dibuang melalui sebuah
proses yang disebut RNA
splicing
-
RNA
splicing
dilakukan oleh RNA-Protein
structure yang disebut Spliceosome

-
Sinyal khusus dari mRNA menunjukkan
dimana keberadaan intron dan exon.
Sinyal-sinyal itu yaitu letak sambungan 5’ dan 3’ dan titik percabangan. RNA splicing akan membuat intron
menjadi loop (simpulan) dan memotongnya dari ujung exon.
-
mRNA yang tidak mengandung intron
membentuk cap di ujung 5' dan dilengkapi dengan sebuah ekor poli(A) membentuk
kompleks dengan protein dn bergerak menembus pori-pori di selubung inti menuju
sitolasma. Di sitoplasma mRNA tersebut bergabung dengan ribosom dan mengarahkan
penggabungan asam amino kedalam protein.
3. Tahap Terminasi
-
Pada eukaryotik pada tahap termination mRNA
akan dimodifikasi.
-
mRNA
disebut
pre-mRNA sebelum
semua modifikasi selesai dilakukan.
-
Ujung 3’ mRNA dibuat menjadi lebih spesifik
sehingga dapat dikenali oleh sejumlah protein RNA binding dan yang memodifikasi
RNA yang akan berikatan RNA
polymerase II.
-
CstF
(Cleavage stimulation factor F) and CPSF (Cleavage and polyadenylation
specificity factor), berikatan dengan RNA polymerase II, memotong RNA pada ujung
3’ dan
bantuan enzim lain yaitu poly-A
polymerase, menambah nukleotida adenosine (A) untuk membuat ekor –A (poly-A-tail).
Dapat diringkas bahwa
RNA eukariotik mengalami transkripsi di inti oleh tiga RNA polimerase yang
berbeda. Transkrip primer mengalami modifikasi dan pemangkasan untuk
menghasilkan RNA matang yang kemudian berpindah ke sitoplasma untuk ikut serta
dalam proses translasi. Prekursor mRNA mengalami pemprosesan yang paling
ekstensif. Prekursor mRNA memiliki sebuah “cap” yang ditambahkan di ujung 5'
dan sebuah “ekor” poli (A) di ujung 3'. Pada prekursor mRNA, ekson daerah yang
membentuk mRNA matang dipisahkan dari intron, daerah yang tidak memiliki fungsi
mengkode dan dikeluarkan oleh reaksi penyambungan. Selama reaksi penyambungan,
ekson-ekson saling dihubungkan untuk menghasilkan mRNA matang. Pada eukariot,
prekursor tRNA dan rRNA juga mengalalami modifikasi dan pemangkasan walaupun
tidak seluas seperti mRNA.
A. Proses Transkripsi pada Bakteri (Prokariot)
Sel bakteri hanya
mempunyai satu RNA polimerase. Pengikatan RNA polmerase ke regio promotor pada
DNA menyebabkan untai-untai DNA terbuka dan terpisah di dalam suatu regio yang
panjang. Sewaktu polimerase melakukan transkripsi DNA, regio heliks yang belum
ditranskripsi terpisah, sementara regio cetakan DNA yang telah ditranskripsi
kembali bergabung dengan pasangannya. Faktor sigma dibebaskan sewaktu rantai
RNA yang sedang tumbuh mencapai panjang sekitar 10 nukleotida. Reaksi
pemanjangan terus berlangsung sampai RNA polimerase menjumpai sinyal terminasi
(penghentian) transkripsi. Salah satu jenis terminasi terdiri dari pembentukan
lengkung tajam di transkrip, mendahului sejumlah residu U. Jenis kedua mekanisme
terminasi meliputi pengikatan suatu protein , faktor rho yang menyebabkan
pelepasan transkrip RNA dari cetakan.
Sistron adalah regio
pada DNA yang mengkode sebuah protein. Pada bakteri, mRNA biasanya dibentuk
dari sebuah operon sebagai suatu transkrip polisistonik (transkrip yang
mengandung informasi untuk menghasilkan sejumlah protein yang berbeda).
Transkrip polisistonik ditranslasikan selagi mengalami transkripsi. Transkripsi
ini tidak mengalami modifikasi atau pemangkasan dan tidak mengandung intron.
Beberapa protein yang berbeda dihasilkan selama translasi transkrip
polisistonik, satu untuk masing-masings sistron.
Dapat
disingkat bahwa pada bakteri sebuah RNA polimerase menghasilkan prekursor mRNA,
rRNA, dan tRNA. Karena bakteri tidak memiliki inti, ribosom berikatan dengan
mRNA sewaktu sedang ditranskripsikan dan sintesis protein berlangsung bersamaan
dengan proses transkripsi.
3.
PROSES TRANSLASI
Setelah tahap
transkripsi yaitu transkripsi DNA
menjadi RNA dimana informasi diubah dari bahasa DNA menjadi bahasa RNA (A,T,G,C menjadi A,U,G,C). Selanjutnya tahap translasi
dimana penterjemahaan bahasa RNA (A,U,G,C)
diubah
menjadi protein (Asam
amino).
Informasi
pada mRNA dibaca dalam kelompok dari 3 nukleotida. Kelompok 3 nukleotida itu disebut
kodon. Urutan kodon pada mRNA mengatur urutan asam amino pada protein. Urutan DNA à urutan RNA = urutan kodon à urutan asam amino = protein.
RNA tersusun dari 4 macam nukleotida – A, U, G and C. Sebanyak 64 kombinasi
kodon yang mungkin yang dapat terbentuk dari keempat nukleotida tersebut, tapi hanya
ada 20 asam amino yang biasanya ditemukan pada protein sehingga satu asam amino
dapat dikode oleh lebih dari satu kodon. Kodon dan asam amino yang dikode
disebut kode genetik. Asam-asam amino dihubungkan oleh ikatan peptida. Protein biasanya mengandung
ratusan asam amino (polipeptida). mRNA dibaca dalam kelompok tiga
nukleotida sehingga akan ada tiga
reading frame dimana
mRNA
dapat dibaca.
Translasi adalah proses penterjemahan informasi di mRNA
menjadi asam amino. Mesin penterjemah di dalam sel meliputi ribosom dan RNA transfer (tRNA) tRNA adalah penyadur yang melekat ke asam amino
tertentu. tRNA mengandung
urutan basa tertentu yang disebut anti kodon, yang melekat ke mRNA melalui
pasangan basa yang sesuai. Anti kodon yang dibawa oleh tRNA sesuai dengan kodon
yang terdapat pada mRNA
yang mengkode asam amino tertentu. Perlekatan asam amino dengan tRNA yang
sesuai diatur oleh enzim amino-acyl
tRNA synthetases. Tiap asam amino memiliki enzim amino-acyl tRNA synthetasenya
sendiri. Asam amino dilekatkan ke ujung 3’ tRNA dengan ikatan yang berenergi tinggi. Energi yang
tersimpan pada ikatan ini akan digunakan dalam proses sintesis protein
nantinya. Ikatan asam amino dengan tRNA yang sesuai dipastikan melalui mekanisme
proof reading yang
dilakukan oleh amino-acyl
tRNA synthetase.
Ribosome merupakan
protein yang dibentuk asam-asam amino melalui ikatan peptida. Ribosome adalah struktur protein-RNA yang dibuat antara ribosomal RNA and protein. Ribosome meliputi dua sub unit
yang tidak sama ukurannya.Sub unit yang lebih kecil bertugas untuk memastikan
bahwa kodon pada mRNA berpasangan secara tepat dengan anti kodon. Sub unit yang
lebih besar berperan dalam pembentukan ikatan peptida antara dua asam amino.
Kedua sub unit akan bersama hanya pada saat translasi.
Tiga
tahapan pada translasi:
1.
Inisiasi : Pada tahap ini translation machinery melekat pada
mRNA.
2.
Elongasi : Pada tahap ini translation machinery bergerak di
sepanjang mRNA dan
mensintesis polipeptida berdasarkan urutan basa pada
mRNA.
3.
Terminasi
: Ketika mRNA selesai dibaca dan protein telah disintesis, maka translation machinery melepaskan diri
dari mRNA.
1. Inisiasi
Untuk memastikan
bahwa mRNA dibaca dengan baik, maka translasi selalui dimulai pada kodon AUG yang disebut dengan initiation codon atau start kodon. Asam
amino yang melekat pada kodon ini adalah methionin (Met/M) dan tRNA yang memulai
translasi disebut initiator
tRNA.
Initiator tRNA merupakan
Met-tRNA
khusus yang berbeda dari Met-tRNA
lainnya. Inisiasi pada translasi difasilitasi oleh protein yang dikenal dengan initiation factors (IF).
Tahap 1: tRNA
initiator dimuat ke dalam unit kecil ribosom dengan bantuan IF (initiation factors).
Tahap 2: Tahap 2: Unit kecil ribosom
melekat pada 5’ cap
(Eukaryotes) /Shine Delgarno sequence (Prokaryotes)
mRNA dan bergerak di sepanjang rantai mRNA sampai menemukan kodon AUG
Tahap 3: Setelah kodon
AUG pertama ditemukan, maka IF melepaskan diri dan unit besar ribosom melekat
pada unit kecil ribosom.

1. Elongasi
Ribosom
bergerak di sepanjang rantai mRNA dan menghubungkan asam amino menurut codon
yang sesuai.
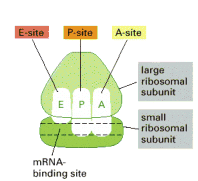
Ribosom
memiliki 3 tempat khusus yaitu E,
P dan A.
-
A
(Amino Acyl Site): tempat dimana tRNA yang baru masuk
melekat.
-
P
(Peptidyl site): tempat dimana ikatan peptida yang baru
dibentuk.
-
E
(Exit site): tempat dimana tRNA yang telah kosong meninggalkan
ribosom.
1. Terminasi
Terminasi
translasi ditandai dengan adanya kodon khusus yang disebut stop codon (UAA, UAG, UGA). Tidak ada tRNA dengan
anti kodon yang dapat mengenali stop kodon. Ketika A site ribosom menemukan stop kodon, hal ini
menyebabkan perlekatan protein yang disebut release factor. Perlekatan ini
menyebabkan pelepasan polipeptida dari ribosom. Setelah polipeptida melepaskan
diri dari ribosom, maka ribosom memisah.
Perbedaan
pada proses translasi prokariot dan eukariot dapat dilihat pada proses inisiasi.
Inisiasi
Eukariot
Pada
eukariot inisiasi translasi terdiri dari pembentukan kompleks yang terdiri dari
metionin tRNA, mRNA, dan sebuah ribosom. Metionin tRNA (Met-tRNA) mula-mula
membentuk kompleks dengan suatu faktor inisiasi (faktor inisiasi eukariotik 2
(elF2) dan GTP. Kompleks ini kemudian mengikat subunit ribosom kecil (40 S).
Cap pada ujung 5' mRNA mengikat faktor inisiasi (elF4E) yang dikenal sebagai cap binding protein (CBP). Kemudian
beberapa elF ikut bergabung, dan mRNA kemudian berikatan dengan kompleks
40S-Met-tRNA. Dalam suatu reaksi yang memerlukan hidrolisis ATP, subunit
ribosom kecil melakukan scan terhadap mRNA sampai kodon AUG pertama ditemukan. elF
lain terikat, GTP mengalami hidrolisis dan faktor inisiasi dibebaskan, dan
subunit ribosom besar (60 S) terikat. Ribosom sekarang menjadi lengkap. Ribosom
ini mengandung satu subunit kecil dan subunit besar. Terdapat dua tempat
pengikatan untuk tRNA, yang dikenal sebagai tempat P (peptidi) dan A
(aminoasil) pada ribosom. Selama inisiasi Met-tRNA berikatan dengan tempat P.
Inisiasi Prokariot
Inisiasi
prokariot dan eukariot berbeda. Pada bakteri metionin-tRNA yang sedang dalam
proses inisiasi mengalami formilasi menghasilkan formil-metionin-tRNA yang ikut
serta dalam pembentukan kompleks inisiasi. Pada prokariot diperlukan hanya tiga
IF (inisiation factor) untuk menghasilkan kompleks ini, sedangkan eukariot
memerlukan selusin atau lebih eIF. Ribosom juga berbeda ukurannya. Prokariot
memiliki ribosom 70S yang terdiri dari subunit 30S dan 50S, sedangkan eukariot
memiliki ribosom 80S yang terdiri dari subunit 40S dan 60S. mRNA bakteri tidak
memiliki cap. Identifikasi triplet AUG untuk inisiasi pada prokariot terjasi
sebagai konsekuensi pengikatan sebuah urutan (yang dikenal sebagai urutan
Shinr-Dalgamo) pada mRNA dengan urutan komplementer dekat ujung 3' rRNA 16S
pada subunit ribosom kecil.




Tidak ada komentar:
Posting Komentar